
Testikler
De testiklene [ Hodn ] eller (mer sjelden) den eller Hode [ hoːdə ] (via mellomhøytysk Hode og gammelhøitysk Hodo av Indo - * skeu (t) eller "cover, cover") testikkelen (fra latin testiculus , diminutiv av testis "vitne [om virilitet ], testikkel", flertall: testikler ; gammelgresk ὄρχις orchis ), teknisk også testis og testiculus , er et sammenkoblet, indre mannlig kjønnsorgan av mange seksuelt reproduserende vevdyr . I likhet med eggstokkene til kvinnelige individer tilhører den kjønnsorganene ( gonader ) og produserer sædfilamentene ( sædceller ). I tillegg dannes mannlige kjønnshormoner ( androgener ), spesielt testosteron , i testiklene . Hos virveldyr dannes testiklene embryonalt i bukhulen, men hos de fleste pattedyr vandrer de inn i pungen ( pungen ).

anatomi
Pattedyr
Størrelse og beliggenhet

Menneskets testikkel er grovt plommeformet , veier ca 20 gram og har et gjennomsnittlig volum på 20-25 ml. Gjennomsnittlig lengde er 5 cm, tykkelsen ca 3 cm. Testiklene utvikler seg ikke til full størrelse før puberteten og når sin maksimale størrelse i det 4. tiåret av livet. Med alderen avtar testikkelstørrelsen igjen. Testikkelvolumet gir blant annet informasjon om funksjonens tilstand. Hvis testikkelvolumet er under 8 ml, kan det antas at sædproduksjonen bare virker i begrenset grad eller ikke i det hele tatt. På den annen side produseres testosteron fremdeles opp til et volum på 1,5 ml; under det er testikkelen vanligvis dysfunksjonell.
Hos pattedyr varierer testikkelformen fra avrundet til eggformet. Det er klare forskjeller i størrelse, men det er ikke noe nært forhold til kroppsvekt. De største testiklene i dyreverdenen har sørlige hval , med hver 500 kg utgjør de 2% av kroppsvekten. Gnagere , sauer (opptil 300 g hver) og tamsvin (opptil 750 g hver) har relativt store testikler, mens rovdyr er relativt små . Hos dyr med en sesongmessig reproduksjonsperiode er testikkelstørrelsen også utsatt for sesongmessige svingninger, testiklene er betydelig større i paringssesongen enn i hvileperioden.
I de fleste pattedyr er begge testiklene hos kjønnsmodne individer i pungen ( pungen ) eller pungenlignende hudposer. Testiklene oppstår i bukhulen , men vandrer gjennom lyskekanalen inn i pungen rundt fødselstidspunktet, hos gnagere ikke før puberteten . Denne prosessen kalles testikulær nedstigning ( Descensus testis ). Hos noen pattedyr ( f.eks. Hamstere , flaggermus ) er det en sesongavstigning av testiklene, og testiklene er bare utenfor bukhulen i paringssesongen. Innen pattedyr er det imidlertid noen grupper av dyr der testiklene vanligvis forblir i bukhulen, den såkalte testiconda . Testiklene kan forbli på stedet for systemet (som hos elefanter ) eller de kan stige ned, men henger fortsatt i bukhulen (for eksempel hos hvaler , se også tabell).
| Testiconda | ||
| Ingen testikkel nedstigning | Ufullstendig nedstigning av testikkel | Sesongbasert testikulær nedstigning |
| Monotremes , gullmoldvarper , spissmus , Igeltenreks , hyraxes , elefanter , sjø kyr , tre-toed sloth , anteaters | Hvaler , armadillos | Moles , snegler , aardvarks , spissmus , flaggermus , noen gnagere |
Anatomisk struktur
Den eksterne anatomiske strukturen til testikelen er basert på epididymis som grenser til og smelter sammen med den . Testikelseksjonen som peker mot hodet på epididymis kalles hodende ( Extremitas capitata ), seksjonen som peker mot halen på epididymis kalles halen ( Extremitas caudata ). På slutten av halen er det ofte en funksjonsløs, vorteformet rudiment av den såkalte Müllers kanal , som er kjent som testikkelvedhengen ( vedlegg testis , et Morgagni-hydatid ). Kanten mot epididymis er kanten av epididymis ( Margo epididymalis ), overfor den er den frie kanten ( Margo liber ). I tillegg kan det skilles mellom en overflate som peker mot sentrum ( Facies medialis ) og en overflate som peker utover ( Facies lateralis ).
De testiklene stige inn i en bule i peritoneum og den indre stammen fascia (referert til her som den fascia spermatica interna ), idet prosessen vaginalis . Prosessen med skjeden er en av testikkelhylsene inne i pungen. Peritoneum-delen av denne fremspringet kalles vaginal hud ( tunica vaginalis testis ). Den strekker innsiden av pungen (såkalt veggark , lamina parietalis eller periorchium ), og snur seg deretter inn som en dobbelt lamella og dekker testiklene som et tarmark ( lamina visceralis eller epiorchium ). Det er et veldig smalt mellomrom mellom de to bladene, cavum vaginale , som sørger for at testikelen kan bevege seg i pungen. Forbindelsespunktet mellom de to bladene er testikkel CrosuS ( mesorchium ), som brukes til å feste testikkel til pungen. Testikelen er også i halen med en kort stripe koblet til epididymis (testis eget bånd, ligament testis proprium ). Dette fortsetter fra epididymal halen som et epididymal tail ligament ( ligamentum caudae epididymidis ) og fester i tillegg testiklene indirekte til gulvet i pungen. Pungen løftemuskelen ( Musculus cremaster ) fester seg også til skjeden, da et beskyttelsesapparat trekker testiklene nærmere bukveggen når den er berørt eller kald. Hos gnagere og pattedyr med sesongavstamning, sjelden også hos enkeltpersoner fra andre pattedyr, kan muskelen trekke testikkelen helt tilbake i bukhulen (" pendulum testicle ").
En tykk, hvitaktig bindevevskapsel , tunica albuginea, ligger rett under bukhinnen som dekker testikelen . Det sikrer organets mekaniske styrke og opprettholder et visst indre trykk. Fra denne kapsel septa trekke inn i det indre og dele testiklene i testikulære lobules ( lobules testis ). Mannens testikkel har omtrent 350 testikulære knopper. Septaen danner også en kropp av bindevev, mediastinum testis , som i menneskelig anatomi også kalles corpus highmori .
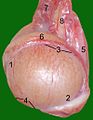
Testikler, epididymis og spermatisk ledning hos en hannkatt :
1 hodeende
2 haleende
3
kant av epididymis 4 fri kant
5 testikkel mesenteri
6 epididymis
7 pleksus av testikkelarterie og vene
8 spermatisk kanal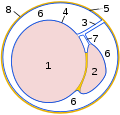
Tverrsnitt gjennom vaginalprosessen:
1 testikkel
2 epididymis
3
testikkel mesenteri 4 organark av vaginal membran
( epiorchium )
5 veggark av vaginal hud
( periorchium )
6 cavum vaginale
7 epididymal mesentery
8 intern spermatisk fascia
Seksjon gjennom en oksens testikkel :
(blodkar injisert med rød gelatin )
1 testikkel parenkym
2 mediastinum testis
3 tunica albuginea
4 epididymis hale
5 epididymis hode
6 sædstreng med
tendril kronglet av testikkelarterien



Fartøy og nerver
.jpg)
Testikelen forsynes med blod via testikkelarterien ( testikkelarterien ). Det oppstår, tilsvarende plasseringen av den embryonale feste av testikelen (se nedenfor), rett bak nyrearterien, direkte fra abdominal aorta i korsryggen. Hos dyr med testikulær avstamming må testikkelarterien forlenges tilsvarende og løper langs baksiden av bukveggen i en kort mesenteri ( mesorchium proksimal ) til lyskekanalen . Utenfor bukhulen kommer den inn i sædlederen . Her ligger den i tette spiralviklinger, den såkalte tendril convolute . Hos okser er for eksempel en to meter lang arterieseksjon innkvartert på en spermatisk ledningslengde på 13 cm. Tendril-bunten er pakket inn i tendril plexus ( Plexus pampiniformis ) i testikkelvenen ( Vena testicularis ). Dette skaper et stort kontaktområde mellom innkommende og utgående blod, som fungerer som en varmeveksler . Temperaturen i pungen er noen grader under den indre kroppstemperaturen, noe som er viktig for dannelsen av fruktbar sædceller hos pattedyr med testikulær nedstigning. Det innkommende blodet i arterien blir avkjølt av dette arrangementet ved blodstrømmen fra venen.
Testikkelarterien går langs kanten av epididymis til enden av halen på testikelen. Derfra beveger den seg inne i testikkelkapslen i den frie kanten hos de fleste pattedyr ( drøvtyggere , for eksempel, gjør et unntak ) tilbake til hodeenden. Dens grener slynger seg i tunica albuginea over sideflatene og går over testikulær septa inn til mediastinum testis og derfra sentrifugalt tilbake til seminiferous tubules, rundt hvilke de danner et kapillærnettverk .
Den innervasjon av testikkelen medieres av de sympatiske del av det autonome nervesystemet . De nervefibre kommer fra stammen av korsryggen og, innpakning rundt testiklene arterien som en flette ( testiklene plexus , synonym: overlegen spermatica nerve ), strekker seg til testiklene. En andre gruppe nervefibre løper fra tverrganglier i stammen med sædkanalen ( ductus deferens ) til testiklene ( plexus deferentialis , syn. Nervus spermaticus inferior ). De efferente nervefibrene når hovedsakelig blodårene og regulerer dermed blodstrømmen og temperaturen i testikelen. Deltakelse i fin kontroll av sæddannelse , sædtransport og hormonproduksjon i testiklene til noen pattedyr blir for tiden diskutert, men denne kontrollen utføres først og fremst via hormoner . De cellelegemer av disse nervefibre som bærer informasjonen til den sentrale nervesystemet ( visceroafferences ) er plassert i den dorsale ganglia korsryggen. De overfører smerteopplevelser (tarmsmerter), men en stor del av testiklens høye følsomhet overfor smerte overføres via de følsomme nervefibrene i testikkelhylsene (grener av kjønnsnervenerven ). Den høye følsomheten for berøringsstimuli gjør testiklene og pungen til en erogen sone . På den annen side utnyttes den store følsomheten for smerte også i BDSM ( kuk og ball tortur ) og tortur ; Klemming av testiklene kan føre til sjokk . En nevralgi av genitofemoral nerve, som kan forekomme, for eksempel etter kirurgisk korreksjon av en lyskebrokk , manifesterer seg i testikelsmerter.
De lymfekar av testikkel løpe sammen med testikler venene til de lumbale lymfeknuter ( Lymphonodi lumbales ) bakover av aorta, i hjemlige pattedyr også til de iliac lymfeknutene ( Lymphonodi iliaci mediales ) ved aorta-gren. I testikkelkreft kan metastaser forekomme i disse lymfeknuter , som ligger i det retroperitoneale rommet i magen .
Gjenværende akkordater
I alle andre akkordater ligger testiklene i kroppshulen og under nyrene.
I den skulderløse er det ingen kompakt testikkel, gonadene er fortsatt segmentert, i Asymmetron er bare den rette testikelen til stede. I hagfish er den langstrakte testikelen også bare dannet på den ene siden i form av et flikete bånd. De runde munnene har sammenkoblede testikler som strekker seg over hele kroppshulen.
Innenfor bruskfisken , bare i kragehaien , den anatomisk og morfologisk mest primitive haien, utvikler hele testikelsystemet seg til en langstrakt gonade, i de andre representantene, som i de andre virveldyrene , bare midtpartiet (se nedenfor). Testiklene til de virkelige haiene er også langstrakte, og strålene er korte og flate. I dogfish og elektriske stråler er testiklene langt foran, nær perikardiet , ellers i den midtre eller bakre bagasjeromsdelen. I de fleste benfisk er testiklene langstrakte. I ekte benfisk er de plassert under nyrene og svømmeblæren og er festet via mesorchium. Hos noen abborarter smelter begge testiklene sammen i bakenden. Den tynne tunica albuginea pigmenteres av og til i benfisk . Samtidig kan eggstokkvev vises i testiklene til fisk (hermafroditt, ovotestis ). Sagabbor og brasme er hermafroditter , så de har både testikler og eggstokker og kan endre sex i løpet av livet.
Amfibier enten har langstrakt ( haleamfibier , krypende amfibier ) eller avrundet ( frosk amfibier ) testiklene. De er festet til stammeveggen eller urnal nyre via mesorchium . En distinkt fettlegeme dannes medialt (i retning av midtplanet ) av testikelen. I salamandere smelter flere testikulære avdelinger sammen i en lap, antall avdelinger øker med alderen. Hos amfibier begynner urinveiene og sædene å skille seg. De epididymale kanalene ( ductuli epididymidis ) åpner seg bare inn i Wolff-kanalen kort før cloaca . Mannlige padder er hermafroditter. Foran testiklene er budorganet , en primitiv eggstokk, som utvikler seg fra den fremre gonadale anlage .
I reptiler er testiklene foran og under nyrene i nærheten av binyrene . Testiklene er ovale, mer avrundet i skilpadder og langstrakte i slanger . Epididymis er mediale (mot midten av kroppen) i den respektive testikelen. Bindevævsrammen til testikelen er dårlig utviklet.
Også hos fugler ligger testiklene foran nyrene på binyrene. I motsetning til eggstokken, som bare er ensidig hos fugler, er testene alltid paret. Testikkelstørrelsen viser de største sesongmessige svingningene i virveldyrene; den øker for eksempel hos passerinefugler med opptil 800 ganger i løpet av paringssesongen og avtar igjen under paringsdvalen. I et hus drakes testiklene er i paringstiden ca 8 cm lang og 4,5 cm bred. Bindevævsstrukturen til testikelen er bare litt utviklet hos fugler, tunica albuginea er tynn, og det er ingen mediastinum testis . Den endoskopiske undersøkelsen av testikkelen spilte tidligere en viktig rolle i å bestemme kjønnet til arter som ikke viser noen seksuell dimorfisme , men i dag har den i stor grad blitt erstattet av molekylærbiologiske metoder.
Hvirvelløse dyr
Asexual og seksuell reproduksjon forekommer i cnidarians . Under seksuell reproduksjon frigjøres kimcellene i det omkringliggende vannet eller mageområdet ved å sprekke epitelet . Ribmaneter er vanligvis hermafroditter og har testikler og eggstokker innebygd i mesogloea under de kammelignende blodplatene ("ribbeina") .
Primal munn (protostomia)
De fleste flatormer (blodplater) er hermafroditter. De har ikke et kroppshulrom ( coelom ), testiklene ligger ved siden av eggstokkene i et bindevevsområde ( interstitium ) i dyret, når det gjelder bendelorm i hvert bendelormlem ( proglottid ). Noen flatorm ( Catenulida og Acoelomorpha ) har ikke kjønnsorganer . Også bukhårede (Gastrotricha) har ikke noe coelom, ferskvannsartene reproduserer aseksuelt via parthenogenese . Saltvannsrepresentantene er hermafroditter, men bare kjønnsorganene av ett kjønn er funksjonelt aktive. Kimcellene frigjøres via sædbunker . Rotatorier (Rotatoria) har en pseudocoeloma med parvise eller uparede testiklene, i enkelte arter såkalte dverg hanner opptrer i andre ikke hanner, tar gjengivelse deretter sted via jomfru generasjon. Cycliophora reproduserer aseksuelt i det såkalte fôringsstadiet. I seksuell reproduksjon forekommer dverghanner. De har to eksterne "testikler" og et kopulatorisk organ ved siden av limskiven. Når det gjelder ripeormer (Acanthocephala), løper et såkalt kjønnsbånd gjennom kroppen, på den nedre tredjedelen av de to testiklene. En sædkanal går fra hver testikkel til penis.
Priapulida og korsettdyr (Loricifera) har sammenkoblet testikler som er forenet med protonephridia for å danne et urin- og seksuelt apparat . I rundorm (Nematoda) er det både separate kjønnarter og hermafroditter. Den langstrakte testikelen ligger under tarmen og åpner seg også inn i den (se fig.). String ormer (taglormer) har paret testiklene i pseudocoeloma. Tardigrades (Tardigrada) er separate kjønn. Mens kroppshulrommet representerer et pseudokoelom, er det en ekte coelom rundt de uparede testiklene. Colibus (Onychophora) har sammenkoblet testikler som åpner seg i en vanlig vas deferens via kanaler. I leddyrene (Arthropoda: tusenben , insekter , krepsdyr og chelicerater ) har hannene paret testikler i den blodfylte pseudocoel ( hemocoel ) i magen eller de bakre stammesegmentene . Det er også hermafroditter i noen leddyr.
De leddormer har koblet testiklene. Belteorm er for det meste hermafroditter, men gjødsler hverandre. Testiklene er lokalisert i coelom , i meitemark i det 10. og 11. kroppssegmentet, og vasedferensene åpner utover i det 15. segmentet. Multi-bust , derimot, er for det meste unisexual og mannlige representanter har testikler i hvert segment av kroppen. I bløtdyr (Mollusca) er det hermafroditter, hermafrodittkirtler ( Ovotestis ) og separate kjønnsformer. Coelom reduseres til to hulrom rundt gonadene ( gonadocoel ) og rundt hjertet ( pericardium ) bak den . I armpods (Brachiopoda) ligger testiklene i coelom (nærmere bestemt i metacoel ), kjønnsceller er avledet via metanephridia .
Neumünder (Deuterostomia)
I pigghuder (Echinodermata) har havhvaler bare en testikkel, i kråkeboller og havhvaler fyller testiklene nesten hele metacoel, i sjøliljer (tre til fem gonader) og sjøstjerner (to per arm) de ligge i armene og åpne mellom dem Armer med en kjønnsåpning hver.

Innvendige organer i en benfisk:
7 testikler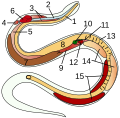
Innvendige organer i en slange:
14 testikler
Sted for en huskuk:
(unntatt fordøyelsesorganer)
3 testikler
Rundorm:
5 testikler
Blueprint av en pod med gonader i metacoel




Fin struktur og funksjon hos virveldyr
Testikkellappene inneholder hver to til fire spiralformede seminiferous tubuli ( tubuli seminiferi contorti s. Convoluti ), som representerer testikkelens parenkym . De er omtrent 50 til 80 cm lange og 150 til 300 um i diameter. Veggen består av et bindevevshjell med kontraktile myofibroblaster , en kjellermembran og kimepitel ( epithelium spermatogenicum ).
Dette epitelet består av sædceller eller kimceller ( Cellulae spermatogenicae ) og Sertoli-celler . Sædcellene dannes fra kimcellene ( spermatogenese ). Siden spermatogenese er testikkelens viktigste oppgave, er kimcellene også hyppigst tilstede i testiklene når det gjelder mengde. Under sæddannelse transporteres de påfølgende utviklingsstadiene til kimcellene ( spermatogonia → spermatocytter → spermatider → sæd) gradvis mot lumen . Sæddannelse tar mellom 35 (mus, gris) og 64 dager (menneske), men da er ytterligere modning i epididymis nødvendig slik at sædcellen kan befruktes. Dette varer en uke hos de fleste pattedyr og 8 til 17 dager hos mennesker. Hos menn frigjøres rundt 200 til 300 millioner sædceller fra epididymis per utløsning . Ved hyppigere utløsning reduseres mengden sædceller fordi den daglige sædproduksjonskapasiteten er begrenset. Det avhenger av testikkelmassen og antall Sertoli-celler og er mellom 45 og 200 millioner sædceller per dag hos menn.
Den andre viktige komponenten i seminiferous tubuli er Sertoli-celler ( Epitheliocyti sustentantes ). De er omtrent 70 til 80 µm lange og løper radialt gjennom hele germinalepitelet opp til lumen. Sertoli-cellene har en støttende og ammende funksjon for sædcellene, de nærer sædcellene, sørger for at de har et riktig hormonelt miljø og bruker plasmabevegelser for å transportere dem til lumen. I tillegg degenererte Sertoli- celler fagocytose sædceller og celleavfall som oppstår under sædutvikling. Sertoli-cellene aktiveres av det follikkelstimulerende hormonet i (FSH) hypofysekontrollert , dets fordeling på dannelsen av hormoninhibitor B- påvirkning. I tillegg utskiller de det androgenbindende proteinet , det anti- Müllerian- hormonet og en kaliumrik sædvæske.
Sertoli-cellene har mange prosesser som omgir kimcellene. Disse celleprosessene forbinder seg ved basen av seminiferous tubuli via tette kryss med de nærliggende Sertoli-celler og danner dermed den såkalte blodtestikulære barrieren . Dette begrepet er faktisk misvisende, fordi denne barrieren ikke ligger mellom blod og testikkelvev, men løper mellom spermatogonia og spermatocytter, og deler derfor testikkelrørene på en sirkulær måte i et basalt og et lumenvendt (adluminal) rom . Blodtestikkelbarrieren er ugjennomtrengelig for de fleste proteiner og beskytter sædceller fra mutagener og fra kroppens eget forsvar . Sistnevnte er nødvendig fordi de første sædcellene først dukker opp etter at lymfocyttene er blitt innprentet (se selvtoleranse ), så immunforsvaret vil anse dem for å være fremmede celler. Imidlertid spiller også antiinflammatoriske cytokiner , som antagelig er androgenavhengig , og cellene i immunsystemet i testiklene ( dendritiske celler , makrofager ) også en rolle i beskyttelsen mot autoimmune reaksjoner .
I mange virveldyr smelter de kronglete seminiferous tubuli sammen til en kort rett seminiferous tubule ( tubulus seminifer rectus ) i begge ender . De rette tubuli er foret av et enkeltlagsepitel og åpner seg inn i et kanalsystem i mediastinum, testikkelnettverket ( rete testis ). Kanalsystemet i testikkelnettverket er også for det meste foret av et enkeltlagsepitel (to-lags hos storfe). Hos menn, hingster og gnagere er testikkelnettverket imidlertid overveiende på periferien til testiklene ("extratesticular rete"). Flere slyngende ductuli efferentes testis løper fra testikkelnettverket inn i epididymal hodet og forenes der for å danne epididymal kanal. Hos pattedyr er det omtrent 15 efferent duktler, antallet varierer innen virveldyr mellom en (f.eks. Stråler ) og 32 (f.eks. Axolotl ).
Vevet mellom de kronglede seminiferous tubuli kalles interstitium . I de fleste virveldyr utgjør det rundt 10 til 20% av testikkelvevet, i ekstreme tilfeller som woodchuck nesten 70%. I tillegg til bindevev, blodkar og nervefibre inneholder interstitium også Leydig-celler ( Endocrinocyti interstitiales ). Via spesielle cellekontakter ( gap junctions ) danner de sammenkoblede celleforeninger, såkalt funksjonell syncytia . Avhengig av det luteiniserende hormonet (LH), produserer Leydig-cellene mannlige kjønnshormoner (androgener som testosteron og androstanolon ), så vel som oksytocin , som fremmer motiliteten til seminiferøse tubuli. Testikelen er dermed også et endokrine organ . Testosteron får spermatidene til å modnes i seminiferous tubuli. For å komme gjennom blod-testikkelbarrieren til dens virkningssted, trenger den det androgenbindende proteinet fra Sertoli-cellene. Androgenene har også en rekke effekter i kroppen, inkludert å fremme utvikling av sekundære seksuelle egenskaper , ha en anabole effekt og kontrollere seksuell atferd . I tillegg produserer Leydig-cellene en rekke andre hormonelt aktive peptider som virker på naboceller ( parakrin ) eller på selve Leydig-cellen ( autokrin ) som utgjør dem .

Histologisk bilde av testikulært parenkym hos en villsvin:
1 lumen av en kronglete seminiferous tubule
2 spermatider
3 spermatocytter
4 spermatogonia
5 Sertoli celler
6 myofibroblaster
7 Leydig celler
8 kapillærer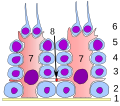
Kimepitel i seminiferous tubule :
1 kjellermembran
2 spermatogony
3 spermatocyte 1. ordre
4 spermatocyte 2. ordre
5 spermatids
6 modne spermatids
7 Sertoli celle
8 tett kryss
(blod-testikkelbarriere)
Skjema for den indre strukturen i testikelen og epididymis :
1 tunica albuginea
2 septa
3 testikellapper
4 mediastinum testis
5 kronglete seminiferous tubules
6 rette seminiferous tubules
7 rete testis
8 ductuli efferentes testis
9 epididymis
10 begynnelsen av vas deferens



Hormonell kontroll

Den hormonell kontroll av testis utføres ved rykk og napp ved nerveceller i median forhøyningen i hypothalamus dannede gonadoliberin (GnRH). GnRH virker ikke direkte på testiklene, men stimulerer dannelsen av hormonene LH og FSH i den fremre hypofysen . Frigjøringen av disse hormonene styres også av testiklene i seg selv via en negativ tilbakemeldingsmekanisme: FSH-sekresjon inhiberes av inhibin B produsert av Sertoli-celler, mens GnRH-sekresjon inhiberes av testosteron produsert av Leydig-celler . De sesongmessige svingningene i testisens størrelse og aktivitet hos mange dyr medieres av undertrykkelse av GnRH-sekresjon under hvile under påvirkning av dagslyslengden. Den eksakte mekanismen er ennå ikke kjent i detalj: Hos pattedyr er antagelig involvert opioider , dopaminerge nevroner og melatonin i denne kontrollsløyfen, hos fugler også skjoldbruskkjertelhormonene .
LH binder seg til en membranreseptor av Leydig-cellene og induserer således syntesen av androgener . I dette tilfellet er kolesterol gradvis, blant annet pregnenolon og progesteron , omdannet til testosteron, ved hjelp av to forskjellige syntetiske veier (Δ4 og Δ5) mulig. LH-effekten på Leydig-cellene forsterkes prolaktin , men med en overproduksjon av prolaktin oppstår ved nedregulering av LH-reseptorer for inhibering av testosteronsyntese. LH kan også indusere dannelse av androgener i binyrebarken ; dehydroepiandrosteronet som dannes der når testiklene via blodet og kan brukes der som en testosteronforløper. Cirka 97% av androgener dannes i testiklene (ca. 7 mg / dag hos menn), den gjenværende delen i binyrene . Androgener virker på germinalepitelet og blir bundet til et protein, og blir også transportert til sine andre målorganer via blodet.
FSH binder seg til de tilsvarende reseptorene på Sertoli-cellene. Både FSH og testosteron kontrollerer spermogenese. FSH setter i gang spermiogenese, testosteron fremmer mitotisk og meiotisk celledeling og dermed dannelsen av spermatocytter fra spermatogonia, mens FSH igjen fører til den endelige modningen av spermatider til sæd.
Undertrykkelsen av den hormonelle stimuleringen av testikkelfunksjonen blir for tiden intensivt undersøkt i utviklingen av prevensjonsmidler for menn . Testosteron eller dets kombinasjon med GnRH-antagonister eller gestagener som progestin anses for tiden å være de mest lovende kandidatene. De fører til sterkt reduserte testosteronkonsentrasjoner i testikelen og dermed til en sterk eller fullstendig reduksjon i sæddannelsen. I veterinærmedisin har et preparat basert på GnRH-analogen Deslorelin ( Suprelorin ® ) blitt godkjent siden 2008 , som undertrykker fruktbarheten hos menn i flere måneder. I tillegg er en vaksine for griser ( Improvac ® ) godkjent, noe som fører til dannelse av antistoffer mot GnRH og dermed undertrykker testikkelfunksjonen.
Utviklingshistorie

Forutsetningen for seksuell reproduksjon er separasjonen av cellene som er spesialisert for reproduksjon (kimceller) fra de normale kroppscellene (somatiske celler). Denne separasjonen er allerede utført i ciliater i form av en mikrokjerne , tydeligere enn i sfæriske alger , der hovedcellegruppen av kroppens celler er i motsetning til en liten gruppe kimceller ( gonidia ), som imidlertid ennå ikke er utviklet i form av et avgrenset organ . Tilstedeværelsen av testikler (eller i prinsippet kjønnsorganer) er ikke en grunnleggende egenskap ved flercellede celler.
I tilfelle bilateria vises en tredje kimblad , mesoderm , og dermed komplekse organer , for første gang . Imidlertid er seksuell reproduksjon i mange virvelløse dyr fortsatt kombinert med muligheten for aseksuell reproduksjon. Det er ofte en generasjonsendring her , dvs. den seksuelle følger en aseksuell reproduktiv syklus.
Differensiering av kjønnsorganene i testikler og eggstokker er et kjennetegn på segregerte arter. Så langt er det ikke avklart om hermafroditisme eller kjønnssegregering er den plesiomorfe egenskapen til bilateria. Begge kjønnsorganene kommer fra det samme systemet i embryonal utvikling. Til tross for denne kjønnsskillingen er reproduksjon uten befruktning ( parthenogenese ) mulig i mange dyregrupper , som kan sees på som en redusert form for seksuell reproduksjon. Hanndyr vises bare her i unntakstilfeller. Parthenogenese finnes i mange taxa , fra rotorer til noen øgler . Opp til amfibiene kan hybridformer eller en endring i kjønn ( dikogami ) også bli funnet under ontogenese . Det er mulig å omdanne eggstokkene til testikler ( proterogyny ) så vel som testiklene til eggstokkene ( proterandry ).
Hos de fleste dyr bestemmes det genetisk om den opprinnelig kjønnsfordelingen av kjønnsorganene resulterer i en testikkel eller en eggstokk . I ormer og fluer bestemmes kjønnet av forholdet mellom X-kromosomer og autosomer . I tilstandsdannende insekter oppstår testikler i avkommet fra ufruktede egg og eggstokkene hos dyr med et diploid sett med kromosomer, dvs. fra befruktede egg. Hos pattedyr bestemmes sex av Y-kromosomet . På dette kjønnskromosomet ( gonosom ) er et gen ( kjønnsbestemmende region av Y , Sry ) lokalisert, som samhandler med gener fra andre kromosomer, og (hos mennesker fra 7. uke etter befruktning) fører til dannelsen av testikkelbestemmende faktor . Dette initierer utviklingen av testiklene og dermed det mannlige kjønn generelt. Den koder for en rekke transkripsjonsfaktorer , de såkalte HMG-proteiner ( høy mobilitetsgruppeproteiner ). Disse proteinene har mange andre funksjoner i andre vev, og de nøyaktige mekanismene som er involvert i testikkeldannelsen blir for tiden intensivt undersøkt. Med uttrykket av Sry skiller Sertoli-cellene seg ut, som blant annet produserer det anti-mulleriske hormonet og dermed forårsaker regresjon av de mulleriske kanalene . Den videre utviklingen av testikelen og de andre egenskapene til det mannlige kjønnet styres av androgener. For noen dyregrupper bestemmes imidlertid kjønnet av miljøfaktorer. I tilfelle noen amfibier og mange reptiler ( skilpadder , alligatorer ) , for eksempel, avhenger kjønnet av inkubasjonstemperaturen. ( Se også. Temperaturavhengig kjønnsbestemmelse )
Hos mange dyr er kjønnsorganene utviklet i nært forhold til utskillelsessystemet ( primitiv nyre , nefridia ), spesielt urinveiene deles som et system som kan sammenlignes, og det er grunnen til at begge organsystemer i virveldyr som urin- og seksuelle apparater oppsummerer. Hos virvelløse dyr er imidlertid opprinnelsesstedet, plasseringen og kanalene veldig forskjellige, slik at man antar at seksuell reproduksjon har oppstått flere ganger og uavhengig av hverandre i evolusjonen . Kompleksiteten til kjønnsorganene avhenger ikke av evolusjonsstadiet; det er for eksempel veldig høyt i flatorm.
Vertebrat embryologi
Testikkelutvikling hos virveldyr
|

I embryoet oppstår testiklene og eggstokkene fra det samme systemet , den såkalte kjønnsryggen . Den dannes i området av urinal nyre og strekker seg opprinnelig fra thorax til lenden . Hos de fleste virveldyr blir bare den midterste delen av dette langstrakte systemet selve gonaden, de gjenværende delene utvikler seg til kjønnsbåndene . Under påvirkning av Sry vandrer de primordiale kimcellene fra plommesekken inn i gonadal anlage (hos mennesker i den 6. embryonale uken) og epitelet i det primitive kroppshulen ( coelom ) vokser fingerlignende inn i systemet som såkalt primære kimlinjer .
Kimen eller testikkelledningene trenger gjennom kjønnsorganet og vokser rundt de primære kimcellene. Kirtlene deles midlertidig inn i cortex og marg, selv om det i mannlige embryoer bare margen utvikler seg til testikelen, mens cortex trekkes tilbake. Lignende prosesser finner sted i genetisk kvinnelige individer, men senere dannes eggstokken fra hjernebarken mens margen degenererer. Forbindelsen mellom testikelsnorene og overflaten går til slutt tapt. Fra testikkelsnorene, via de Sry-uttrykkende pre-Sertoli-celler, utvikler Sertoli-cellene seg, som regnes som arrangørene av videre testikkelutvikling og derved samhandler med myoidcellene . Spermatogonia oppstår fra de primære kjønnscellene .
Inni danner testikkelkablene et nettverk av sammenkoblede ledninger, det senere testikulære nettverket ( rete testis ). Testikkelnettverket kobles til noen urnale tubuli som blir de ledende kanalene til epididymis- hodet . Urinkanalen ( Wolff duct ) brukes også som en epididymal kanal og spermatisk kanal som en spermadreneringsbane. De lumen av de sædkanaler utvikles ikke før puberteten , i amfibier etter metamorfose , inntil da de testikulære ledningene er fast.
Tunica albuginea , rammeverket av bindevev i testikkelen og Leydig-celler oppstår fra den mesodermale delen av testikkelanus . Leydig-cellene kan også bli funnet i den tidlige testikkelutviklingen; de uttrykker steroidogen faktor 1 (Sf1), og deres testosteronproduksjon bestemmer signifikant utviklingen av de mannlige kjønnsorganene.
Aldring
Hos menn - i motsetning til kvinner ( se overgangsalderen ) - er det ikke plutselig opphør av funksjonen til gonadene i en definert alder. Både produksjon av hormoner og modning av kimcellene blir potensielt bevart i alderdommen. Faktisk er farskap dokumentert frem til det tiende tiåret av livet . I virkeligheten er det imidlertid markerte individuelle forskjeller, og mange menn blir før eller siden infertile , noe som ikke skal forveksles med impotens . De nøyaktige årsakene til disse forskjellene er ikke kjent i detalj; blant annet mistenkes vaskulære faktorer. Fra et statistisk synspunkt, fra rundt det fjerde tiåret av livet og utover, blir testiklene involvert, som utvikler seg veldig sakte, med en reduksjon i vekt, størrelse og sædproduksjon. For den enkelte er det imidlertid knapt noen spådommer å komme.
De strukturelle endringene er derfor også gjenstand for et bredt spekter, men et blandet bilde av normale og tydelig atrofiske testikulære tubuli kan sees på som typisk . Et markert fall i testosteronproduksjonen observert hos noen menn kan føre til Climacterium viril med hetetokter , hodepine og andre symptomer .
Utforskningshistorie

I antikken og middelalderen ble testikkelen bare ansett som en transittstasjon for sæd. Alkmaion von Kroton (tidlig på 5. århundre f.Kr.) mistenkte hjernen som opprinnelsen til sæd, som når testiklene via blodkar . De atomists ( Anaxagoras , Demokrits ) og Aristoteles inngår i ryggmargen i denne banen, Galenus (125-199) mistanke om opprinnelsen av spermieceller i blodkarene de når testiklene gjennom hvilke. Disse ideene fortsatte i middelalderen. Leonardo da Vincis anatomiske tegninger viser forbindelser mellom testikkel og lunger og hjerne, fordi da Vinci mistenkte opprinnelsen til den "åndelige kraften" til sæd i hjernen, mens testiklene bare gir det materielle grunnlaget for de "nedre impulser".
Strukturell utforskning
Det var først med begynnelsen av opplysningstiden på 1600-tallet at ideene om mannlig sæd ble avmystifisert og den direkte forbindelsen mellom testikler og reproduksjon anerkjent. Den første moderne beskrivelsen av strukturen til testikelen kommer fra Reinier de Graaf (1641–1673). Nathaniel Highmore beskrev i 1651 testikkelens bindevevskropp ( Corpus Highmori ), som ble oppkalt i 1830 av Astley Paston Cooper Mediastinum testis . I 1677 oppdaget oppfinneren av mikroskopet , Antoni van Leeuwenhoek , sædceller, som han mente var miniatyriserte, forhåndsdannede organismer ("seedyr").

Med utviklingen av histologiske teknikker, kunne også den fine strukturen i testikkelen belyses. I 1841 anerkjente den sveitsiske anatom Albert von Koelliker den direkte forbindelsen mellom testikulære tubuli og sædceller for første gang og oppdaget at sædcellene i disse tubuli oppstår som produkter av cellulær differensiering. I 1850 beskrev Köllikers elev Franz von Leydig mellomcellene ( Leydig-celler ) for første gang .
I 1865 oppdaget Enrico Sertoli støttecellene ( Sertoli-celler ). I 1871 lyktes Victor Ebner i å skille Sertoli-cellene fra spermatogonia, og fem år senere laget La Valette St. George begrepet "spermatogonia" og klassifiseringen av de individuelle utviklingstrinnene til sædcellene, noe som fremdeles er vanlig i dag. Sertoli-cellene ble lenge sett på som syncitia , det var ikke før 1956 at Don W. Fawcett og Mario H. Burgos var i stand til å bevise at hver Sertoli-celle har sine egne cellegrenser.
Hugo Ribbert anerkjente allerede i 1904 at karmin som ble administrert i blodet ikke kom inn i lumen av seminiferous tubuli og testikkelnettverket. I lang tid ble denne oppdagelsen forsømt, selv om det var det første beviset på blod-testikkelbarrieren. Denne kunnskapen ble først tatt opp på slutten av 1950-tallet, og i 1963 lyktes J. Brökelmann å bevise de tette kryssene til Sertoli-cellene som det morfologiske grunnlaget for blod-testikkelbarrieren. Paul J. Gardner og Edward A. Holyoke var i stand til å belyse den fine strukturen i blod-testikkelbarrieren et år senere.
Hormoner
Selv om effektene av kastrering hadde vært kjent i tusenvis av år, det var ikke før 1849 at Arnold Adolph Berthold lyktes i eksperimentelt demonstrerer dannelsen av hormoner i testiklene ved hjelp av testikkel transplantasjoner i kuker. I en veldig gammel alder foretok Charles-Édouard Brown-Séquard selveksperimenter på slutten av 1800-tallet med væske fra testene til hunder og marsvin, som han tilskrev en foryngende og styrkende kraft, selv om det var mer homøopatiske hormonmengder. som han oppnådde på denne måten. På begynnelsen av 1900-tallet ble transplantasjon av dyre testikler under bukveggen betraktet som et middel til foryngelse, spesielt wiener Robert Lichtenstern og Eugen Steinach var hovedpersoner i denne metoden. Steinach ønsket å oppnå denne foryngelsesprosessen ved å ligere vas deferens (hans mest berømte pasient var Sigmund Freud ) og beskrev testikkeltransplantasjon som en "terapi" for homofili . Etter 1945 gikk disse kontroversielle fremmedbildene av moten.
I 1903 publiserte Pol Bouin og Paul Ancel først kunnskapen om at Leydig-cellene er stedet der manlige kjønnshormoner produseres. I 1931 isolerte Adolf Butenandt og Kurt Tscherning androsteron (en metabolitt av testosteron) fra urinen til menn, i 1935 var Ernst Laqueur i stand til å isolere testosteron selv fra oksetestikler og laget også navnet på dette hormonet (fra testikler "testikler" og "testikler") steroid ").
Eksistensen av ikke-steroide hormoner i testiklene ble postulert så tidlig som på 1920-tallet, men det var først i 1932 at D. Roy McCullagh demonstrerte sin funksjonalitet i kastrerte rotter og kalte det inhibin . Selv om bioassays for dette hormonet ble utviklet på 1960-tallet , var dets eksistens kontroversiell i flere tiår og ble ikke allment akseptert før 1979. I 1984/85 ble hormonets struktur og undertyper avslørt. Med avklaring av DNA- sekvensen som koder for hormonet , ble tilknytningen av inhibinen til gruppen av β-transformerende vekstfaktorer anerkjent i 1985 .
Mens forholdet mellom LH og testosteron allerede var kjent på 1960-tallet, ble FSH-avhengigheten til Sertoli-celler først bevist i 1984 av Joanne M. Orth .
Kjønnsdifferensiering
Det kromosomale grunnlaget for kjønnsdifferensiering ble allerede ryddet mellom 1910 og 1916, først og fremst gjennom arbeidet til Thomas Hunt Morgan på fruktfluer , som han mottok 1933 Nobelprisen i fysiologi eller medisin . Alfred Jost anerkjente i 1947 at kjønnsorganene primært bestemmes av det kvinnelige kjønnet og at det mannlige kjønnet er avhengig av testosteron. Imidlertid var det først på begynnelsen av 1960-tallet at Y-kromosomet ble identifisert som en viktig faktor hos pattedyr. Den nøyaktige plasseringen av genet for den testikkelbestemmende faktoren ble ikke bestemt før i 1990; dets forskjellige funksjoner er ennå ikke kjent i detalj og er gjenstand for aktuell forskning.
Utviklingsforstyrrelser og sykdommer
Skader på testikelen forekommer hos mennesker som stump traumer , spesielt i kampsport og kamper. Det er en risiko for blødning under testikkelkapslen ( hematocele ), som vanligvis krever kirurgisk behandling. Skader ved åpning av pungen (punktering og impaling sår, hos dyr også biter, piggtråd, etc.) kan forårsake testikkelbetennelse (se nedenfor) eller til og med abscesser , samt peritonitt på grunn av den åpne forbindelsen av vaginal prosess til bukhulen .
Misdannelser
En anorki er fraværet av begge testiklene. Hvis bare en testikkel dannes, snakker man om et monarki . Omtrent 5% av mannlige barn som opereres for manglende nedstigning av testikler har bare en eller ingen testikkel. De har ofte små bindevevsknuter med spredte Leydig-celler. Siden en funksjonelt intakt testikkel er helt nødvendig for det mannlige kjønnsuttrykket, må minst en intakt testikkel ha vært til stede i den embryonale fasen, som kan ha gått tilbake senere.
I sjeldne tilfeller, som et resultat av utviklingsforstyrrelser, kan eggstokker også vises i tillegg til testikler hos mennesker og andre pattedyr ( hermafroditisme verus - "ekte" hermafroditter; se også interseksualitet ). Med visse misdannelser i gonader ( gonadal dysgenese ), blir testiklene ikke opprettet, forblir underutviklet eller inneholder eggstokkvev ( ovotestis ).
En svært sjelden uønsket utvikling er det splenogonadal fusjon med en forbindelse mellom testikler og miltvev .
Med forstyrrelser av den migrering av testikkel ( Maldescensus testis ) forskjellige posisjonsmessige avvik kan forekomme. Testikelen kan forbli i bukhulen ( kryptorchidism , "abdominal testicle "), bli sittende fast i lyskekanalen ("inguinal testicle", "glide bein") eller ta feil rute og for eksempel komme til å ligge under huden av lysken eller det indre låret ( testikkel ektopi ). Maldescensus testis er en av de vanligste misdannelsene hos mennesker og forekommer hos 3 til 5% av nyfødte og 33% av premature babyer; kryptorchids forekommer også med en lignende frekvens hos husdyr og fører til ekskludering fra avl. Hvis testikelen er feil plassert, kan det ikke dannes fruktbar sæd på grunn av temperaturfølsomheten i kimepitelet, men androgenproduksjonen beholdes. Unnlatelse av å synke ned i mer enn to år kan føre til tap av spermatogonia og dermed irreversible endringer i testikelen.
Arvelig kort vekst av testiklene ( testikkelhypoplasi ) er relativt vanlig hos kjæledyr. Testikulær hypoplasi kan også være forårsaket av kromosomale lidelser ( Klinefelter syndrom ), infeksjoner eller hormonelle lidelser .
En unormal utvidelse av testiklene kalles makroorchidi .
Betennelse i testiklene
Betennelse i testiklene ( orchitis ) kan oppstå med skader på pungen med invasjon av bakterier eller med noen smittsomme sykdommer. Orchitis er en mulig komplikasjon av kusma , Coxsackie-virusinfeksjoner og vannkopper hos mennesker . Også brucellose og tuberkulose kan manifestere seg på testiklene. Hos dyr også tuberkulose og kan brucellose og pseudotuberculosis ( sau ), hesteinfeksiøs anemi og kattinfeksiøs anemi ( katter ) ledsages av en orkiditt. Betennelse i testiklene kan føre til krymping av testiklene ( testikkelatrofi ) og infertilitet , fordi det ikke kan dannes flere sædceller ( aspermi ) eller ingen fungerende sædceller.
Sirkulasjonsforstyrrelser
Som en varicocele kalles åreknuter lignende utvidelser som spesielt venene til venstre på plexus pampiniformis bekymrer seg i sædstrengen. En varicocele kan føre til begrenset sædproduksjon i testikelen på samme side.
Den ansamling av serøs væske i testikulære mantlene kalles en hydrocele eller vann brudd. I tillegg kan væsken også samles i sædstrengen, som deretter blir referert til som hydrocele funiculi spermatici .
En torsjon er en unormal rotasjon av testis, hvor nevnte spiralformede klemming av sædstrengen og dreneringsårene kan føre til at testis dør. Alvorlig testikeltorsjon er en veldig smertefull nødsituasjon, og det kan forventes permanent skade på testikelen etter bare to timer. Den testis tillegget kan også utvikle en såkalt hydrat torsjon.
Sirkulasjonsforstyrrelser med risiko for å utvikle nekrose observeres også i sykdommer i blodårene som Henoch-Schönlein purpura , endangiitis obliterans og panarteritis nodosa hos mennesker, arteritt hos hester og generelt ved trombose .
Svulster
En testikelsvulst er en patologisk forstørrelse av testikelen. Testikulære svulster kan være godartede eller ondartede .
For det meste ufarlige testikkelforstørrelser er cyster . To forskjellige typer cyster kan utvikles på testikelen. Hydroceles er fremspring fra tunica vaginalis testis som inneholder en klar, ravfarget væske. De er forårsaket av skade eller betennelse. Spermatoceles stammer fra rete testis eller epididymis og inneholder sædceller. Teratomer er for det meste godartede svulster i kjønnscellene.
En veldig sjelden årsak til en masse på testikelen kan være splenogonadal fusjon .
Ondartede testikulære svulster ( testikkelkreft ) er delt inn i degenerasjoner av kimcellene ( germinal testikulære svulster : seminomer ) og ikke- seminomer . Degenerering av kimcellene er den vanligste kreft hos menn i alderen 20 til 40 år og utgjør rundt 90% av alle testikulære svulster. Hovedrisikofaktoren er testikler som ikke har migrert inn i pungen. De resterende 10% skyldes svulst degenerasjon av andre vevsdeler ( Sertoli celle svulst , Leydig celle svulst , ikke-Hodgkin lymfom, etc.).
Feilfunksjoner
I tillegg til de ovennevnte sykdommer , kjemiske substanser som krysser den blod-testisbarrieren så som miljøgifter (f.eks kadmium ), additiver til emballasjemateriale (f.eks ftalater , dietylheksyl-adipat ), noen medikamenter (f.eks furazolidon ) og kan hormoner også forekomme ( se også hormonforstyrrende stoffer ) eller ioniserende stråling kan føre til alvorlig svekkelse av epitelet i seminiferous tubuli. Siden sæddannelse er forbundet med svært høye celledeling ( mitose , meiose ), er kimepitelet spesielt følsomt for celletoksiner. Slike skader kan føre til manifold forandringer opp til fullstendig fravær av sædceller ( se også sperm -analyse ).
Utilstrekkelig produksjon av androgener er kjent som hypogonadisme . Dette kan være medfødt, sekundært til sykdommer i testiklene eller på grunn av mangel på gonadotropin (f.eks. Hypofunksjon i hypofysen , olfaktogenitalt syndrom ).
Abdominal kryptorchidisme hos tamfår ("kidney buck")

Testikulær hypoplasi hos en katt, til høyre den normalt utviklede testikkelen.

Distinkt varicocele til venstre.
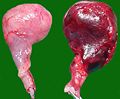
Hund testikler fjernet etter rotasjon (venstre: sunt organ)
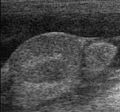
Sonography : hund med abdominal kryptorchidisme og ascites (venstre: testikler, høyre: epididymis)





undersøkelse
Den testiklene undersøkelsen er et viktig grunnleggende undersøkelse for mennesker og dyr med en pung. Dette er hvor testikkelens tilstedeværelse, størrelse, plassering og konsistens kontrolleres. Ultralydundersøkelsen brukes primært som en bildemetode . Testikkelvolumet bestemmes enten ved sammenligning med den såkalte Prader-kjeden eller ved hjelp av ultralydmåling. Diaphanoscopy har fremdeles en viss verdi i diagnosen hydrocele . En testikkel biopsi kan utføres for å ta vevsprøver . Hos dyr med testikler plassert i bukhulen brukes endoskopi primært i tillegg til ultralydundersøkelse .
En funksjonell undersøkelse er opprettelsen av en sædanalyse . Her vurderes sædens antall, form og mobilitet. Bestemmelsen av nivået av inhibin B i blodet brukes som en markør for Sertoli-cellefunksjon og fruktbarhet, men dens informative verdi er kontroversiell.
Leydig-cellestimuleringstesten kan brukes til å oppdage testikkelvev som ikke kan oppdages ved hjelp av bildebehandlingsteknikker .
kastrering

Begrensningen av testikkelfunksjonen kalles kastrasjon. Det kan gjøres ved kirurgisk fjerning av testikkelen ( orkidektomi ), ligering av testikkelkarene ("blodfri kastrasjon"), stråling eller kjemiske stoffer. Kastrasjoner utføres hovedsakelig hos mennesker for testikkelkreft. Kirurgisk fjernede testikler erstattes vanligvis av en testikkelprotese av kosmetiske årsaker .
Som et symbol på disempowerment spiller kastrering også en rolle i mytologien til mange kulturer (se også kastrasjonsfrykt ). I egyptisk mytologi snapper Horus testiklene fra sin motstander Seth . I gresk mytologi fjerner Kronos først testiklene til sin far Uranus og blir senere imponert av sønnen Zeus selv . For å gi avkall på verdslige ønsker, var selve- emasculation av Galloi (prester) i Cybele kult av frygierne , som også spredte seg til antikkens Hellas og Roma, vanlig, slik tilfellet var med hijraene i India . I jødedommen er imidlertid kastrering, både av mennesker og dyr, strengt forbudt. I kristendommen ble kastrering også mislikt. Eunuchs fikk ikke lov til å bli ordinert til prest, men det var strømmer der selvkastrering ble utført som et ritual (se Skopzen ).
Historisk sett ble også slaver , krigsfanger, sangere eller haremsvakter (se Palace Eunuch ) kastrert. Den ikke-medisinsk begrunnede kastrasjonen var spesielt rettet mot forebygging av sekundære seksuelle egenskaper (stemmehøyde, seksuell oppførsel) forårsaket av testosteron. Castrati var populære i det europeiske musikklivet på 1600- og 1700-tallet og ble ofte respektert. Senesino , Farinelli , Caffarelli og Antonio Bernacchi er blant de mest berømte castratiene fra 1700-tallet .
Kastrerte menn kan ikke reprodusere av seg selv. I likhet med geistlige som frivillig hadde avstått , ble de bedømt som mer pålitelige og brukt som funksjonærer og tjenere i forskjellige samfunn. Frivillig kastrering av sexforbrytere er fremdeles en behandlingsmetode, om enn en kontroversiell, i Tyskland og i noen delstater i USA.
I veterinærmedisin brukes kastrasjon, i tillegg til medisinske indikasjoner (testikkelkreft, prostata og analkjertelsykdommer ), fremfor alt for å unngå avkom, for å forbedre håndteringen av kjæledyr ( vallak , okse ), for å øke oppdrettsytelsen og kjøttkvaliteten, hos tamgriser også for å unngå "utsvinelukten" av kjøttet som ble utført. Kastreringer på dyr ble sannsynligvis utført så tidlig som yngre steinalder . Kastrering er en av de få ikke-medisinsk indikerte organfjerningene som fortsatt er tillatt i Tyskland i dag i henhold til dyrevelferdsloven (§ 6), hos svært unge dyr, selv uten å eliminere smerte, som imidlertid ikke er ubestridt.
Kulturell og historisk betydning

I japansk mytologi blir tanuki , demoner ( yōkai ) som ligner på vaskebjørnhunden , ofte representert som et symbol på flaks med store testikler. I det gamle Hellas ble kjønnsorganene til dyr, spesielt okser ( taurobolium ), ofret. Testikler ble ansett som et symbol på virilitet og kreativ styrke. I følge Taylor hadde de en sterkere symbolsk kraft enn penis til slutten av 1500-tallet .
botanikk

Plantene, som ligner mannlige kjønnsorganer i utseendet, ble antatt å ha en afrodisiakum og fruktbarhetsøkende effekt. Den greske filosofen Theophrastus von Eresos kalte dem Orchis , det greske navnet på testikler, på grunn av likheten mellom orkidéens to pærer og testiklene. Forbruket deres sies å bidra til fødselen av en gutt ("orkide"). Orchis ga senere navnet hele orkideefamilien .
Navnet avokado er avledet fra det indiske ordet ahuacatl (testikkel), som refererer til den testikkelignende formen på frukten av dette treet.
I kunst
I kunst, i motsetning til fallus , spiller ikke testikler en sentral rolle utenfor erotikk og pornografi . “Blood and Testicles”, et fonologisk ordspill på blod-og-jord-ideologien , brukes ofte nedsettende i kunstkritikk .
En av karakterene i Thomas Manns Tristan er Mr. Klöterjahn ("Klöten" er det lavtyske uttrykket for testikler, et symbol på egnethet til liv og vitalitet). I romanen Sanningen om Sascha Knisch fra 2002 ( Eng . Tittel The Truth About Sascha Knisch. 2003) av den svenske forfatteren Aris Fioretos , er testiklene ledemotivet . Den tyske filmen Eierdiebe tar for seg emnet testikkelkreft og tap av testikkel.

Våpenskjoldet til det italienske adelshuset Colleoni fra Bergamo viser flere par testikler og er sannsynligvis en hentydning til coglione , et italiensk navn på testikler. En statue av Bartolomeo Colleoni med dette våpenskjoldet på basen ber hovedpersonen i første bind av Heinrich Manns tredelte roman The Goddesses or The Duchess of Assy's Three Novels .
Som mat

Testikler blir behandlet som mat i mange regioner. Tyskland var det eneste landet i EU hvor testikler ble forbudt som mat. I henhold til EUs regulativ nr. 853/2004 av 29. april 2004 er testikler de eneste kjønnsorganene som mat, alle andre er klassifisert som uegnet til konsum ( konfiskert ).
Diverse
- Den amerikanske Gregg Miller mottok Ig Nobelprisen for medisin i 2005 for å utvikle testikkelproteser for kastrerte hunder .
- I mars 2006 hadde Volkswagen annonsert Golf GTI på plakater med "Turbo Cojones" i USA . På engelsk står begrepet cojones for mot og dristighet, på spansk, der cojones står for "testikler" og i daglig tale "skjæring" eller "mot", betyr ordkombinasjonen bokstavelig talt "turbo testikler". Kampanjen ble trukket tilbake etter protester.
- De poseformede kjertlene (høysekkene) under beverens kjønnsben var også kjent som "testikler" tidligere.
litteratur
- AJP van den Brock: Gonader og måter å utføre. I: Bolk et al. (Red.): Håndbok for den komparative anatomien til virveldyrene . Bind 6, Urban & Schwarzenberg, Berlin 1933, s. 1–154.
- W. Busch, A. Holzmann (red.): Veterinær andrologi. Schattauer, Stuttgart 2001, ISBN 3-7945-1955-8 .
- U. Gille: Mannlige reproduktive organer. I: F.-V. Salomon et al. (Red.): Anatomi for veterinærmedisin . Enke, Stuttgart 2004, ISBN 3-8304-1007-7 , s. 389-403.
- R. Hautmann, H. Huland: urologi . Springer, Berlin 2006, ISBN 3-540-29923-8 .
- H.-G. Liebich: Funksjonell histologi hos tampattedyr. 4. utgave. Schattauer, Stuttgart 2003, ISBN 3-7945-2311-3 .
- JD Neill (red.): Knobil og Neills reproduksjonsfysiologi. 3. Utgave. Academic Press, Amsterdam 2005, ISBN 0-12-515400-3 .
- PE Petrides: Endokrine funksjoner IV. Hypothalamus-hypofysesystem og målvev. I: G. Löffler, PE Petrides (red.): Biokjemi og patobiokjemi. 7. utgave, Springer, Berlin 2003, ISBN 3-540-42295-1 , s. 865-908.
- U.N. Riede et al.: Mannlig kjønnssystem. I: U.-N. Riede et al. (Red.): Generell og spesiell patologi . Thieme, Stuttgart 1989, ISBN 3-13-683302-3 , s. 768-779.
- B. Vié: testikler. Fête de paires, mytologi, les undertøy, d'une curiosité culinaire, les attributs du sujet, lexique. Edition de l'Epure, Paris 2005, ISBN 2-914480-58-X (mange oppskrifter, beriket med kulturell og historisk informasjon).
- R. Wehner, W. Gehring: Zoologi. 23. utgave, Thieme, Stuttgart 1995, ISBN 3-13-367423-4 .
- U. Welsch: Sobotta lærebokhistologi. Urban & Fischer, München 2002, ISBN 3-437-42420-3 .
weblenker
Individuelle bevis
- ↑ Walther Graumann: Kompakt lærebokanatomi. Volum 3, Schattauer, Stuttgart 2004, ISBN 3-7945-2063-7 , s. 265.
- ↑ H. Sosnik: Studier på størrelsen på menneskelig mannlig gonade i biomorfose, alkoholforgiftning og skrumplever - en gjennomgang og egne funn. I: Gegenbaurs morfologiske årbok. Vol. 134, nr. 5, 1988, s. 733-761, PMID 3224804 .
- ^ David AE Spalding: Whales of the West Coast . Harbor Publishing, 1999, ISBN 1-55017-199-2 .
- ↑ I. Gerendai u a.: Innervering og serotoninerge reseptorer i testis interagerer med lokal virkning av interleukin-1beta på steroidogenese. I: Autonomic Neuroscience. 7. juli 2006, PMID 16829209 .
- ↑ I. Ducic, AL Dellon: Testikulær smerte etter reparasjon av lyskebrokk: en tilnærming til reseksjon av kjønnsgrenen i kjønnsorganisk nerve. I: Journal of the American College of Surgeons. Volum 198, nr. 2, februar 2004, s. 181-184, PMID 14759772 .
- ↑ FD Brown et al.: Buders organ i padden Bufo marinus: Effekter av orkidektomi på morfologien og uttrykket av lamina-assosiert polypeptid 2. I: Utviklingsvekst og differensiering. Volum 44, nr. 6, desember 2002, s. 527-535, PMID 12492511 .
- ↑ CF Farias: Buders organ av Bufo ictericus: en lys- og elektronmikroskopi-analyse. I: Micron. Volum 33, nr. 7-8, 2002, s. 673-679, PMID 12475564 .
- ↑ G. Michel: Sex system. I: F.-V. Salomon (red.): Lærebok for fjærfeanatomi . Fischer-Verlag, Stuttgart 1993, ISBN 3-334-60403-9 , s. 197-226.
- ^ RP Amann, SS Howards: Daglig spermatozoal produksjon og epididymal spermatozoal reserver av den menneskelige hannen. I: Journal of Urology . Vol. 124, nr. 2, august 1980, s. 211-215, PMID 6772801 .
- Peters C. Petersen, O. Soder: Sertoli-cellen - et hormonelt mål og 'super' sykepleier for kimceller som bestemmer testikkelstørrelsen. I: Hormone Research Volume 66, No. 4, 2006, s. 153-161, PMID 16804315 (fulltekst) .
- ↑ M. Fijak, A. Meinhardt: testis i immun privilegium. I: Immunologiske anmeldelser . Volum 213, oktober 2006, s. 66-81, PMID 16972897 .
- S RS Swerdloff et al.: Undertrykkelse av spermatogenese hos mennesker indusert av Nal-Glu gonadotropinfrigivende hormonantagonist og testosteron enanthat (TE) opprettholdes av TE alene. I: Journal of Clinical Endocrinology and Metabolism . Volum 83, nr. 10, oktober 1998, s. 3527-3533, PMID 9768659 .
- Matth KL Matthiesson: Effekter av testosteron og levonorgestrel kombinert med en 5alfa-reduktasehemmer eller gonadotropinfrigivende hormonantagonist på spermatogenese og intrestikulære steroidnivåer hos normale menn. I: Journal of Clinical Endocrinology and Metabolism . Vol. 90, nr. 10, oktober 2005, s. 5647-5655, PMID 16030154 .
- ↑ Peter Y. Liu et al. Determinanter av hastighet og grad av spermatogenese Suppression under hormonell prevensjon for hann-: En integrert analyse. I: Journal of Clinical Endocrinology and Metabolism. Volum 93, nr. 5, s. 1774-1783, doi: 10.1210 / jc.2007-2768 .
- ↑ Improvac på vetpharm.uzh.ch
- ↑ a b c d C. Tilmann, B. Capel: Cellular and Molecular Pathways Regulating Mammalian Sex Determination. I: Nylig fremgang i hormonforskning. Volum 57, 2002, s. 1-18.
- ↑ C. Dournon et al.: Temperatur kjønnsomslag hos amfibier og reptiler. I: International Journal of Dev Biology. Volum 34, utgave 1. mars 1990, s. 81-92. PMID 2393628
- ^ E. Nieschlag et al.: Reproduksjonsfunksjoner hos unge fedre og bestefedre. I: Journal of Clinical Endocrinology and Metabolism . Vol. 55, 1982, s. 676-681.
- ↑ H. Bürgi, C. Hedinger: Histologiske testikkelendringer i alderdommen. I: Sveitsisk medisinsk ukentlig. Volum 47, 1959, s. 1236-1239.
- ↑ A. von Kölliker: Bidrag til kunnskap om kjønnsrelasjoner og sædvæsken av virvelløse dyr, sammen med et eksperiment på naturen og betydningen av såkalte frø dyr . Berlin 1841.
- H AHJ La Valette St. George: Om opprinnelsen til frølegemene. I: Arkiv for mikroskopisk anatomi. Volum 12, 1876, s. 797-825.
- ^ J. Brökelmann: Fin struktur av kimceller og Sertoli-celler under syklusen av seminiferous epitel i rotte. I: Celleforskning og mikroskopisk anatomi. Vol. 59, 1963, s. 820-850, PMID 14015736 .
- ^ PJ Gardner, EA Holyoke: Fin struktur av den sveitsiske musen. I. Den begrensende membranen, Sertoli-celle, spermatogonia og spermatocytter. I: Anatomical Record . Volum 150, desember 1964, s. 391-404, PMID 14248309 .
- ↑ AA Berthold: Transplantasjon av testiklene. I: Arkiv for anatomi og fysiologi. Fysiologisk avdeling . Volum 16, 1849, s. 42-46.
- ↑ F. Mildenberger: Foryngelse og "helbredelse" av homofili. Eugen Steinach i sin tid. I: Sex Research. Volum 15, 2002, s. 302-322.
- ^ P. Bouin, P. Ancel: Recherches sur les Cellules interstitielles du testicule chez les mammifères. I: Archives de zoologie expérimentale et générale. Serie 4, bind 1, 1903, s. 437-523.
- ↑ K. David et al.: Om krystallinsk hormon fra testikler (testosteron). I: Journal of Physiological Chemistry . Volum 233, 1935, s. 281-282.
- ↑ A. Lampel: Commentary. I: Gjeldende urologi. Volum 35, 2004, s. 6-8. (Fulltekst) ( Memento fra 14. november 2007 i Internet Archive )
- ↑ Axel Wehrend: Nøkkelsymptomer på gynekologi og fødselshjelp hos hunder. Enke, Stuttgart 2010, ISBN 978-3-8304-1076-8 , s.57 .
- ^ A b c G. Taylor: Castration: An Abbrevtered History of Western Manhood. Routledge, 2002, ISBN 0-415-93881-3 .
- ↑ W. Bittorf: blod og testikler . I: Der Spiegel . Nei. 44 , 1976, s. 228-232 ( Online - 25. oktober 1976 ).
- ^ Peter Philipp Riedl: Epok-bilder - kunstnertypologier. Bidrag til tradisjonelle utkast i litteratur og vitenskap fra 1860 til 1930. Vittorio Klostermann, 2005, ISBN 3-465-03410-4 , ( s. 570 ).
- ↑ Forordning (EF) nr. 853/2004 fra Europaparlamentet og Rådet. ( europa.eu )
- ↑ Miriam Jordan: Good Taste Lost In Ad Translation . På: articles.sun-sentinel.com fra 19. mars 2006 (engelsk).
- ↑ 'Turbo-Cojones', eslogan de Volkswagen que molesta en EEUU . På: elmundo.es 21. mars 2006 (spansk).
- ↑ Foto av plakaten. På: secure.flickr.com fra 20. mars 2006.
- ↑ Dieter Lehmann: To medisinske reseptbøker fra 1400-tallet fra Øvre Rhinen . Del I: Tekst og ordliste . Horst Wellm, Pattensen / Han. 1985; nå (= Würzburg medisinsk historisk forskning. Volum 34). Königshausen & Neumann, Würzburg, ISBN 3-921456-63-0 , s. 194.