
Proteinkinaser
| Proteinkinaser | ||
|---|---|---|

| ||
| cAMP-avhengig proteinkinaseheksamer, Sus scrofa i henhold til PDB 1CMK | ||
| Enzymklassifisering | ||
| EC, kategori | 2.7.-.- , kinase | |
| Svarstype | Fosforylering | |
| Underlag | Proteiner | |
| Produkter | Fosfo-proteiner | |
Proteinkinaser er enzymer som katalyserer overføringen av en fosfatgruppe fra en donor (vanligvis ATP ) til sidekjedehydroksy (OH) -gruppen i en aminosyre . Kinaser er derfor fosfo- transferaser .
Forekomst av kinaser
Ifølge Gerard Manning koder rundt 2% av alle gener i det eukaryote genomet for kinaser. De omtrent 518 kinasene kan 30% av alle proteiner i et cellefosforylat . Summen av alle kinaser i en celle kalles også et kinom (basert på -omikk ). En klassifisering basert på sekvenslikheter av kinase-domenene, kjente biologiske funksjoner, viser 10 grupper, 143 familier og 212 underfamilier i kinomer til mennesker, fluer, ormer og gjær.
Funksjonene til kinasene
Den proteinfosforylering er en meget viktig post-translasjonelle kontrollmekanismen i signaltransduksjon av cellen. Dette inkluderer for eksempel regulering av aktiviteten til enzymer eller transkripsjonsfaktorer . Fosforylering kan aktivere / hemme målproteinet eller bestemme dets subcellulære lokalisering gjennom endringer i ladning eller konformasjon. De fysiologiske effektene av disse proteinfosforyleringene avhenger av det spesielle substratet til kinasen.
Proteinkinasedysfunksjon er årsaken til mange sykdommer. Proteinkinaser er derfor attraktive molekylære mål for legemiddelintervensjon og blir intensivt undersøkt av legemiddelindustrien. “Spesifikke” hemmere av proteinkinaser brukes med suksess i kreftterapi (se for eksempel imatinib i behandlingen av kronisk myeloid leukemi ).
Regulering av kinaseaktivitet
På grunn av deres vidtrekkende innflytelse må proteinkinasene i seg selv være strengt regulert. Regulatoriske faktorer inkluderer:
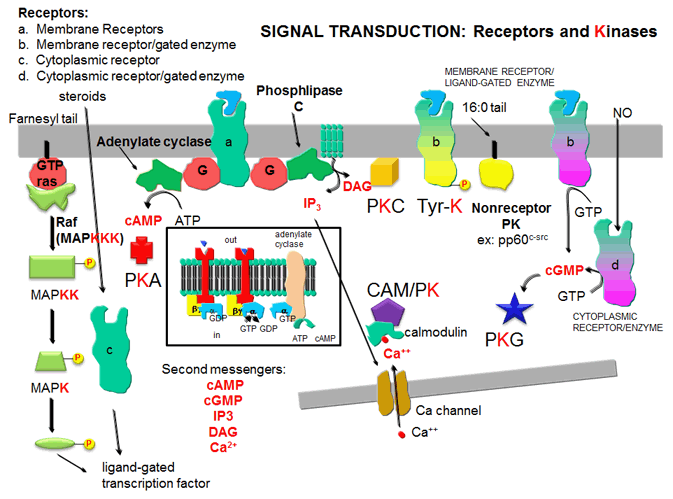
- Kofaktorer / sekundære messenger stoffer ( Ca 2+ , IP 3 , cAMP etc.)
- Aktivator- og inhibitorproteiner
- Pseudosubstrater
- Autoinhibition (del av proteinkinasen peptid- kjede fungerer som en pseudo-substrat)
- Ligandbinding til regulatoriske underenheter
- Fosforylering i det aktive stedet ved
- andre proteinkinaser ( transfosforylering )
- seg selv ( cis -fosforylering / auto-fosforylering)
- Subcellular lokalisering i cellen
Typer proteinkinaser
De fleste proteinkinaser fosforylerer enten aminosyrene serin / treonin eller aromatiske tyrosiner . Imidlertid er det også bispesifikke proteinkinaser ( EC 2.7.12 ; f.eks. MEK eller MAP2K ) som kan fosforylere både serin / treonin- og tyrosinrester.
Serin / treoninkinaser
Disse proteinkinasene ( EC 2.7.11 ) fosforylerer hydroksylgruppene (OH-gruppene) av aminosyrene serin og treonin . Disse kinasene er regulert av:
- cAMP eller cGMP
- 1,2-diacylglyserin (DAG)
- Ca 2+ eller kalmodulin
- PIP 3 og andre fosfolipid- derivater
Det faktum at serin / treoninkinaser ikke fosforylerer alle seriner og treoniner av andre proteiner, bestemmes av interaksjonen med den omkringliggende peptidsekvensen. Disse sekvensene kalles konsensus-sekvenser . På grunn av den lave spesifisiteten fosforylerer ikke disse kinasene individuelle proteiner, men hele proteinfamilier. Disse enzymene inhiberes av det faktum at et pseudosubstrat binder seg til det aktive stedet ved å etterligne målsekvensen til den tilsvarende kinasen, men ikke har serin eller treonin.
Muskelfosforylase kinase A
Dette enzymet ( fosforylasekinase , EC 2.7.11.19 ) er den første Ser / Thr-kinasen som ble oppdaget i 1956. Det er et nøkkelenzym i glykogenmetabolismen .
Proteinkinase A
Dette enzymet har flere funksjoner i cellen. Disse inkluderer regulering av glykogen-, sukker- og lipidmetabolisme . Den proteinkinase A ( EC 2.7.11.11 ) er sammensatt av to domener. Den mindre består av et stort antall β-ark , mens den store underenheten har et stort antall α-helices . Det katalytiske sentrum ligger mellom de to underenhetene. Hvis ATP binder seg til et substrat, vrir underenhetene mot hverandre slik at γ-fosfatgruppen i ATP kommer nær aminosyren som skal fosforyleres, og overføringsreaksjonen kan finne sted.
Du er selv regulert av cAMP. Ved binding av cAMP til en inaktiv tetramer består av to regulerings- og to katalytiske subenheter (R 2 C- 2 ), blir de regulatoriske underenheter atskilt fra de katalytiske de, som muliggjør at fosforylering av andre proteiner. Proteinkinase A er i sin tur regulert av fosforylering.
I tillegg reduserer aktivering av en fosfodiesterase den tilgjengelige mengden cAMP ved å konvertere den til AMP. Som et resultat forårsaker proteinkinase A sin egen inhibering, og derfor kan ingen "permanent aktivering" av kinasen forekomme.
Proteinkinase B
Kinasen, ofte referert til som Akt i den engelskspråklige litteraturen , er et enzym av PI3K / Akt-signalveien som har mange effekter på cellehomeostase og regulerer overlevelse, apoptose , spredning og metabolisme . Akt forekommer i tre nært beslektede isoformer (Akt1, Akt2, Akt3), hvis sekvenser er kodet på henholdsvis kromosomer 14q32, 19q13 og 1q43. Ingen forskjeller i substratspesifisitet kunne bli funnet mellom de enkelte isoformene.
Aktivering skjer i flere trinn. Først rekrutteres fosfatidylinositol-3-kinaser ( PI3K ) til membranen av aktiverte reseptortyrosinkinaser og aktiveres dermed. Aktivert PI3K katalyserer fosforylering av fosfatidylinositol til fosfatidylinositol-3-fosfat eller av fosfatidylinositol-4-fosfat til fosfatidylinositol-3,4-bisfosfat og av fosfatidylinositol-4,5-bisfosfat til fosfatidylinositol-3,4,5-trisfosfat. De resulterende substratene rekrutterer nå proteiner med Pleckstrin Homology (PH) -domener til membranen. Akt som er forankret på denne måten kan nå aktiveres via et ytterligere signalmolekyl som har PH-domener og som fosforylerer fosfoinositidavhengig kinase 1 (PDK1).
Som nevnt ovenfor regulerer proteinkinase B blant annet overlevelse, spredning og dødssyklusen til den berørte cellen. Et eksempel er økt forekomst av glukose reseptor GLUT-4 med et innkommende insulinsignal. Transkripsjonen av denne bæreren fyres opp på en kaskadeaktig måte og inkorporeringen av den initieres av vesikeltransport og innsnevring. Aktiviteten av enzymet kan selv reguleres, for eksempel ved tumor suppressor protein PTEN , som brukes for defosforylering av, for eksempel, fosfatidylinositol-3,4,5-TRIS-fosfat til fosfatidylinositol-4,5-bifosfat (eller andre 3'-fosforylerte fosfoinositider) ansvarlige er. Dette gjør substratet til proteinkinasen uegnet fordi det ikke lenger er i stand til å forankre seg på PH-domenet.
Funksjons- og reguleringsmekanismene er forskjellige og blir fortsatt undersøkt i detalj i dag, ettersom det antas at en mutasjon i visse områder av signalveien (f.eks. PTEN) er et grunnlag for dannelse av svulster .
Proteinkinase C
Betegnelsen proteinkinase C ( EC 2.7.11.13 ) gjelder en familie av proteiner med 12 medlemmer i pattedyr, Ca 2+ , diacylglyserol og en fosfolipid slik. B. trenger fosfatidylkolin for aktivering. I de fleste tilfeller , når vi snakker om disse enzymene , menes proteinkinase Cα .
Proteinkinasene er sterkt konserverte proteiner som består av et N-terminal regulatorisk domene og et C-terminal katalytisk domene. Så lenge enzymet ikke aktiveres av en tumorpromotor som tetradekanoylforbolacetat (TPA) eller en av kofaktorene nevnt ovenfor, er det inaktivt. Den generelle lineære strukturen:
N - pseudosubstrat - TPA / DAG-binding - Ca 2+ binding - ATP-binding - substratbinding - C
Under aktivering beveger proteinkinase C seg til cellemembranen ved hjelp av reseptorer for aktivert proteinkinase C ( RACK-proteiner ). Etter aktivering forblir disse kinasene aktive i lang tid, selv om Ca 2+ konsentrasjonen har redusert igjen. Dette tilskrives virkningen av diacylglyserol , som dannes av fosfatidylinositol av en fosfolipase . Dette aktiveres av de samme signalene som selve proteinkinasen.
Målsekvensen til proteinkinase C er lik den til proteinkinase A ved at den inneholder mange basiske aminosyrerester nær de fosforylerte Ser / Thr-restene. Substrater av proteinkinase C er MARCKS proteiner, MAP-kinaser , transkripsjonsfaktor inhibitor IkB, vitamin D 3 reseptor , raf-kinase , calpain og EGF reseptor .
Tyrosinkinaser
Blant tyrosinkinasene ( EC 2.7.10 ) er det såkalte reseptortyrosinkinaser (RTK) og tyrosinkinaser uten reseptorfunksjon. Rundt 50 forskjellige RTK-er er gruppert i 18 reseptorfamilier . I følge ligandene er disse RTKene gruppert i følgende familier:
Multifunksjonelle kinaser
Noen kinaser fosforylerer metabolitter så vel som andre proteiner. Fosfoglyseratkinase 1 (PGK1) fosforylerer både 3- fosfoglyserat (ved hjelp av ATP) til 1,3-difosfoglyserat og minst 3 proteiner, f.eks. B. Beclin 1 (involvert i autofagi ), PDHK1 (pyruvat metabolisme) og Bcl2 ( apoptose ).
Individuelle bevis
- Ning Manning G. Genomisk oversikt over proteinkinaser. WormBook. 2005 13. des ;: 1-19. Anmeldelse. PMID 18050405
- ↑ Krebs EG, Fischer EH. Fosforylase b til et konverterende enzym av kanin skjelettmuskulatur. Biochim Biophys Acta . 1956 apr; 20 (1): 150-7. PMID 13315361
- ^ Aaronson SA. Vekstfaktorer og kreft. Vitenskap. 1991 22. november; 254 (5035): 1146-53. Anmeldelse. PMID 1659742
- ↑ Zhimin Lu, Tony Hunter: Metabolic Kinases Moonlighting as Protein Kinases . I: Trender i biokjemisk vitenskap . teip 43 , nei 4. april 2018, ISSN 0968-0004 , s. 301-310 , doi : 10.1016 / j.tibs.2018.01.006 , PMID 29463470 , PMC 5879014 (gratis fulltekst) - ( elsevier.com [åpnet 22. juli 2018]).
weblenker
- Reseptorer og kinaser: fem hovedproteinkinaser - representasjon av rollen til store proteinkinaser i signaltransduksjon (engelsk)
- Protein Kinase Resource - Omfattende presentasjon av kinomet med interaktive alternativer for å sammenligne individuelle kinaser med hensyn til funksjon og struktur
{kind=link}